Chloroplasts visible in the cells of Bryum capillare, a type of moss
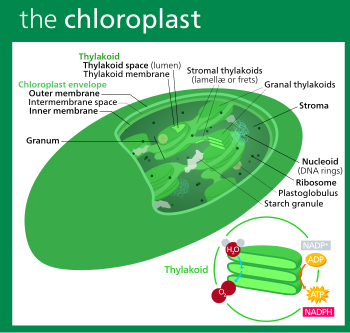
Structure of a typical higher-plant chloroplast
Chloroplasts/ˈklɔːrəˌplæsts, -plɑːsts/[1][2] are organelles that conduct photosynthesis, where the photosynthetic pigment chlorophyll captures the energy from sunlight, converts it, and stores it in the energy-storage molecules ATP and NADPH while freeing oxygen from water in plant and algal cells. They then use the ATP and NADPH to make organic molecules from carbon dioxide in a process known as the Calvin cycle. Chloroplasts carry out a number of other functions, including fatty acid synthesis, much amino acid synthesis, and the immune response in plants. The number of chloroplasts per cell varies from one, in unicellular algae, up to 100 in plants like Arabidopsis and wheat.
A chloroplast is a type of organelle known as a plastid, characterized by its two membranes and a high concentration of chlorophyll. Other plastid types, such as the leucoplast and the chromoplast, contain little chlorophyll and do not carry out photosynthesis.
Chloroplasts are highly dynamic—they circulate and are moved around within plant cells, and occasionally pinch in two to reproduce. Their behavior is strongly influenced by environmental factors like light color and intensity. Chloroplasts, like mitochondria, contain their own DNA, which is thought to be inherited from their ancestor—a photosynthetic cyanobacterium that was engulfed by an early eukaryotic cell. Chloroplasts cannot be made by the plant cell and must be inherited by each daughter cell during cell division.
With one exception (the amoeboid Paulinella chromatophora), all chloroplasts can probably be traced back to a single endosymbiotic event, when a cyanobacterium was engulfed by the eukaryote. Despite this, chloroplasts can be found in an extremely wide set of organisms, some not even directly related to each other—a consequence of many secondary and even tertiary endosymbiotic events.
The word chloroplast is derived from the Greek words chloros (χλωρός), which means green, and plastes (πλάστης), which means "the one who forms".[3]
4.4.1Chloroplast genome reduction and gene transfer
4.4.2Protein synthesis
4.5Protein targeting and import
4.5.1Transport proteins and membrane translocons
5Structure
5.1Outer chloroplast membrane
5.2Intermembrane space and peptidoglycan wall
5.3Inner chloroplast membrane
5.3.1Peripheral reticulum
5.4Stroma
5.4.1Chloroplast ribosomes
5.4.2Plastoglobuli
5.4.3Starch granules
5.4.4RuBisCO
5.5Pyrenoids
5.6Thylakoid system
5.6.1Granal structure
5.6.2Thylakoids
5.6.3Pigments and chloroplast colors
5.6.3.1Chlorophylls
5.6.3.2Carotenoids
5.6.3.3Phycobilins
5.7Specialized chloroplasts in C4 plants
6Location
6.1Distribution in a plant
6.2Cellular location
6.2.1Chloroplast movement
7Function and chemistry
7.1Guard cell chloroplasts
7.2Plant innate immunity
7.3Photosynthesis
7.3.1Light reactions
7.3.1.1Energy carriers
7.3.1.2Photophosphorylation
7.3.1.3NADP+ reduction
7.3.1.4Cyclic photophosphorylation
7.3.2Dark reactions
7.3.2.1Carbon fixation and G3P synthesis
7.3.2.2Sugars and starches
7.3.2.3Photorespiration
7.4pH
7.5Amino acid synthesis
7.6Other nitrogen compounds
7.7Other chemical products
8Differentiation, replication, and inheritance
8.1Plastid interconversion
8.2Chloroplast division
8.2.1Regulation
8.3Chloroplast inheritance
8.3.1Transplastomic plants
9References
10External links
Discovery
The first definitive description of a chloroplast (Chlorophyllkörnen, "grain of chlorophyll") was given by Hugo von Mohl in 1837 as discrete bodies within the green plant cell.[4] In 1883, A. F. W. Schimper would name these bodies as "chloroplastids" (Chloroplastiden).[5] In 1884, Eduard Strasburger adopted the term "chloroplasts" (Chloroplasten).[6][7][8]
Chloroplast lineages and evolution
Chloroplasts are one of many types of organelles in the plant cell. They are considered to have originated from cyanobacteria through endosymbiosis—when a eukaryotic cell engulfed a photosynthesizing cyanobacterium that became a permanent resident in the cell. Mitochondria are thought to have come from a similar event, where an aerobic prokaryote was engulfed.[9] This origin of chloroplasts was first suggested by the Russian biologist Konstantin Mereschkowski in 1905[10] after Andreas Schimper observed in 1883 that chloroplasts closely resemble cyanobacteria.[5] Chloroplasts are only found in plants, algae,[11] and the amoeboid Paulinella chromatophora.
Cyanobacterial ancestor
Main article: Cyanobacteria
Cyanobacteria are considered the ancestors of chloroplasts. They are sometimes called blue-green algae even though they are prokaryotes. They are a diverse phylum of bacteria capable of carrying out photosynthesis, and are gram-negative, meaning that they have two cell membranes. Cyanobacteria also contain a peptidoglycan cell wall, which is thicker than in other gram-negative bacteria, and which is located between their two cell membranes.[12] Like chloroplasts, they have thylakoids within.[13] On the thylakoid membranes are photosynthetic pigments, including chlorophyll a.[14]Phycobilins are also common cyanobacterial pigments, usually organized into hemispherical phycobilisomes attached to the outside of the thylakoid membranes (phycobilins are not shared with all chloroplasts though).[14][15]
Both chloroplasts and cyanobacteria have a double membrane, DNA, ribosomes, and thylakoids. Both the chloroplast and cyanobacterium depicted are idealized versions (the chloroplast is that of a higher plant)—a lot of diversity exists among chloroplasts and cyanobacteria.
Primary endosymbiosis
Primary endosymbiosis A eukaryote with mitochondria engulfed a cyanobacterium in an event of serial primary endosymbiosis, creating a lineage of cells with both organelles.[9] It is important to note that the cyanobacterial endosymbiont already had a double membrane—the phagosomal vacuole-derived membrane was lost.[16]
Somewhere around 1 to 2 billion years ago,[17][18][19] a free-living cyanobacterium entered an early eukaryotic cell, either as food or as an internal parasite,[9] but managed to escape the phagocytic vacuole it was contained in.[14] The two innermost lipid-bilayer membranes[20] that surround all chloroplasts correspond to the outer and inner membranes of the ancestral cyanobacterium's gram negative cell wall,[16][21][22] and not the phagosomal membrane from the host, which was probably lost.[16] The new cellular resident quickly became an advantage, providing food for the eukaryotic host, which allowed it to live within it.[9] Over time, the cyanobacterium was assimilated, and many of its genes were lost or transferred to the nucleus of the host.[23] From genomes that probably originally contained over 3000 genes only about 130 genes remain in the chloroplasts of contemporary plants.[18] Some of its proteins were then synthesized in the cytoplasm of the host cell, and imported back into the chloroplast (formerly the cyanobacterium).[23][24] Separately, somewhere around 500 million years ago, it happened again and led to the amoeboid Paulinella chromatophora.[19]
This event is called endosymbiosis, or "cell living inside another cell with a mutual benefit for both". The external cell is commonly referred to as the host while the internal cell is called the endosymbiont.[9]
Chloroplasts are believed to have arisen after mitochondria, since all eukaryotes contain mitochondria, but not all have chloroplasts.[9][25] This is called serial endosymbiosis—an early eukaryote engulfing the mitochondrion ancestor, and some descendants of it then engulfing the chloroplast ancestor, creating a cell with both chloroplasts and mitochondria.[9]
Whether or not primary chloroplasts came from a single endosymbiotic event, or many independent engulfments across various eukaryotic lineages, has long been debated. It is now generally held that organisms with primary chloroplasts share a single ancestor that took in a cyanobacterium 600–2000 million years ago.[17][19] It has been proposed this bacterium was Gloeomargarita lithophora.[26][27][28] The exception is the amoeboid Paulinella chromatophora, which descends from an ancestor that took in a Prochlorococcus cyanobacterium 90–500 million years ago.[29][28][19]
These chloroplasts, which can be traced back directly to a cyanobacterial ancestor, are known as primary plastids[30] ("plastid" in this context means almost the same thing as chloroplast[9]). All primary chloroplasts belong to one of four chloroplast lineages—the glaucophyte chloroplast lineage, the amoeboid Paulinella chromatophora lineage, the rhodophyte (red algal) chloroplast lineage, or the chloroplastidan (green) chloroplast lineage.[31] The rhodophyte and chloroplastidan lineages are the largest,[16] with chloroplastidan (green) being the one that contains the land plants.[16]
Glaucophyta
See also: Cyanobacteria and Glaucophytes
The alga Cyanophora, a glaucophyte, is thought to be one of the first organisms to contain a chloroplast.[24] The glaucophyte chloroplast group is the smallest of the three primary chloroplast lineages, being found in only 13 species,[16] and is thought to be the one that branched off the earliest.[16][17][32] Glaucophytes have chloroplasts that retain a peptidoglycan wall between their double membranes,[30] like their cyanobacterial parent.[12] For this reason, glaucophyte chloroplasts are also known as 'muroplasts'[30] (besides 'cyanoplasts' or 'cyanelles'). Glaucophyte chloroplasts also contain concentric unstacked thylakoids, which surround a carboxysome – an icosahedral structure that glaucophyte chloroplasts and cyanobacteria keep their carbon fixation enzyme RuBisCO in. The starch that they synthesize collects outside the chloroplast.[14] Like cyanobacteria, glaucophyte and rhodophyte chloroplast thylakoids are studded with light collecting structures called phycobilisomes.[14][30] For these reasons, glaucophyte chloroplasts are considered a primitive intermediate between cyanobacteria and the more evolved chloroplasts in red algae and plants.[30]
Diversity of red algae Clockwise from top left: Bornetia secundiflora, Peyssonnelia squamaria, Cyanidium, Laurencia, Callophyllis laciniata. Red algal chloroplasts are characterized by phycobilin pigments which often give them their reddish color.[33]
Rhodophyceae (red algae)
The rhodophyte, or red algal chloroplast group is another large and diverse chloroplast lineage.[16] Rhodophyte chloroplasts are also called rhodoplasts,[30] literally "red chloroplasts".[34]
Rhodoplasts have a double membrane with an intermembrane space and phycobilin pigments organized into phycobilisomes on the thylakoid membranes, preventing their thylakoids from stacking.[14] Some contain pyrenoids.[30] Rhodoplasts have chlorophyll a and phycobilins[32] for photosynthetic pigments; the phycobilin phycoerytherin is responsible for giving many red algae their distinctive red color.[33] However, since they also contain the blue-green chlorophyll a and other pigments, many are reddish to purple from the combination.[30] The red phycoerytherin pigment is an adaptation to help red algae catch more sunlight in deep water[30]—as such, some red algae that live in shallow water have less phycoerytherin in their rhodoplasts, and can appear more greenish.[33] Rhodoplasts synthesize a form of starch called floridean starch,[30] which collects into granules outside the rhodoplast, in the cytoplasm of the red alga.[14]
Chloroplastida (green algae and plants)
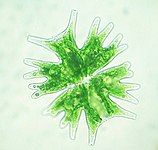
Diversity of green algae Clockwise from top left: Scenedesmus, Micrasterias, Hydrodictyon, Volvox, Stigeoclonium. Green algal chloroplasts are characterized by their pigments chlorophyll a and chlorophyll b which give them their green color.
The chloroplastidan chloroplasts, or green chloroplasts, are another large, highly diverse primary chloroplast lineage. Their host organisms are commonly known as the green algae and land plants.[35] They differ from glaucophyte and red algal chloroplasts in that they have lost their phycobilisomes, and contain chlorophyll b instead.[14] Most green chloroplasts are (obviously) green, though some aren't, like some forms of Hæmatococcus pluvialis, due to accessory pigments that override the chlorophylls' green colors. Chloroplastidan chloroplasts have lost the peptidoglycan wall between their double membrane, leaving an intermembrane space.[14] Some plants seem to have kept the genes for the synthesis of the peptidoglycan layer, though they've been repurposed for use in chloroplast division instead.[36]
Most of the chloroplasts depicted in this article are green chloroplasts.
Green algae and plants keep their starch inside their chloroplasts,[14][32][35] and in plants and some algae, the chloroplast thylakoids are arranged in grana stacks. Some green algal chloroplasts contain a structure called a pyrenoid,[14] which is functionally similar to the glaucophyte carboxysome in that it is where RuBisCO and CO2 are concentrated in the chloroplast.[37]
Transmission electron micrograph of Chlamydomonas reinhardtii, a green alga that contains a pyrenoid surrounded by starch.
Helicosporidium is a genus of nonphotosynthetic parasitic green algae that is thought to contain a vestigial chloroplast.[32] Genes from a chloroplast[38] and nuclear genes indicating the presence of a chloroplast have been found in Helicosporidium[32] even if nobody's seen the chloroplast itself.[32]
Paulinella chromatophora
While most chloroplasts originate from that first set of endosymbiotic events, Paulinella chromatophora is an exception that acquired a photosynthetic cyanobacterial endosymbiont more recently. It is not clear whether that symbiont is closely related to the ancestral chloroplast of other eukaryotes.[16] Being in the early stages of endosymbiosis, Paulinella chromatophora can offer some insights into how chloroplasts evolved.[23][39]Paulinella cells contain one or two sausage shaped blue-green photosynthesizing structures called chromatophores,[23][39] descended from the cyanobacterium Synechococcus. Chromatophores cannot survive outside their host.[23] Chromatophore DNA is about a million base pairs long, containing around 850 protein encoding genes—far less than the three million base pair Synechococcus genome,[23] but much larger than the approximately 150,000 base pair genome of the more assimilated chloroplast.[40][41][42] Chromatophores have transferred much less of their DNA to the nucleus of their host. About 0.3–0.8% of the nuclear DNA in Paulinella is from the chromatophore, compared with 11–14% from the chloroplast in plants.[39]
Secondary and tertiary endosymbiosis
Many other organisms obtained chloroplasts from the primary chloroplast lineages through secondary endosymbiosis—engulfing a red or green alga that contained a chloroplast. These chloroplasts are known as secondary plastids.[30]
While primary chloroplasts have a double membrane from their cyanobacterial ancestor, secondary chloroplasts have additional membranes outside of the original two, as a result of the secondary endosymbiotic event, when a nonphotosynthetic eukaryote engulfed a chloroplast-containing alga but failed to digest it—much like the cyanobacterium at the beginning of this story.[16] The engulfed alga was broken down, leaving only its chloroplast, and sometimes its cell membrane and nucleus, forming a chloroplast with three or four membranes[43]—the two cyanobacterial membranes, sometimes the eaten alga's cell membrane, and the phagosomal vacuole from the host's cell membrane.[16]
Secondary endosymbiosis consisted of a eukaryotic alga being engulfed by another eukaryote, forming a chloroplast with three or four membranes.
Diagram of a four membraned chloroplast containing a nucleomorph.
The genes in the phagocytosed eukaryote's nucleus are often transferred to the secondary host's nucleus.[16]Cryptomonads and chlorarachniophytes retain the phagocytosed eukaryote's nucleus, an object called a nucleomorph,[16] located between the second and third membranes of the chloroplast.[14][24]
All secondary chloroplasts come from green and red algae—no secondary chloroplasts from glaucophytes have been observed, probably because glaucophytes are relatively rare in nature, making them less likely to have been taken up by another eukaryote.[16]
Green algal derived chloroplasts
Green algae have been taken up by the euglenids, chlorarachniophytes, a lineage of dinoflagellates,[32] and possibly the ancestor of the CASH lineage (cryptomonads, alveolates, stramenopiles and haptophytes)[44] in three or four separate engulfments.[45] Many green algal derived chloroplasts contain pyrenoids, but unlike chloroplasts in their green algal ancestors, storage product collects in granules outside the chloroplast.[14]
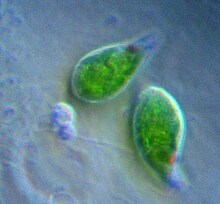
Euglena, a euglenophyte, contains secondary chloroplasts from green algae.
Euglenophytes
Euglenophytes are a group of common flagellated protists that contain chloroplasts derived from a green alga.[16]Euglenophyte chloroplasts have three membranes—it is thought that the membrane of the primary endosymbiont was lost, leaving the cyanobacterial membranes, and the secondary host's phagosomal membrane.[16] Euglenophyte chloroplasts have a pyrenoid and thylakoids stacked in groups of three. Photosynthetic product is stored in the form of paramylon, which is contained in membrane-bound granules in the cytoplasm of the euglenophyte.[14][32]
Chlorarachnion reptans is a chlorarachniophyte. Chlorarachniophytes replaced their original red algal endosymbiont with a green alga.
Chlorarachniophytes
Chlorarachniophytes /ˌklɔːrəˈræknioʊˌfaɪts/ are a rare group of organisms that also contain chloroplasts derived from green algae,[16] though their story is more complicated than that of the euglenophytes. The ancestor of chlorarachniophytes is thought to have been a eukaryote with a red algal derived chloroplast. It is then thought to have lost its first red algal chloroplast, and later engulfed a green alga, giving it its second, green algal derived chloroplast.[32]
Chlorarachniophyte chloroplasts are bounded by four membranes, except near the cell membrane, where the chloroplast membranes fuse into a double membrane.[14] Their thylakoids are arranged in loose stacks of three.[14] Chlorarachniophytes have a form of polysaccharide called chrysolaminarin, which they store in the cytoplasm,[32] often collected around the chloroplast pyrenoid, which bulges into the cytoplasm.[14]
Chlorarachniophyte chloroplasts are notable because the green alga they are derived from has not been completely broken down—its nucleus still persists as a nucleomorph[16] found between the second and third chloroplast membranes[14]—the periplastid space, which corresponds to the green alga's cytoplasm.[32]
Prasinophyte-derived dinophyte chloroplast
Lepidodinium viride and its close relatives are dinophytes (see below) that lost their original peridinin chloroplast and replaced it with a green algal derived chloroplast (more specifically, a prasinophyte).[14][46]Lepidodinium is the only dinophyte that has a chloroplast that's not from the rhodoplast lineage. The chloroplast is surrounded by two membranes and has no nucleomorph—all the nucleomorph genes have been transferred to the dinophyte nucleus.[46] The endosymbiotic event that led to this chloroplast was serial secondary endosymbiosis rather than tertiary endosymbiosis—the endosymbiont was a green alga containing a primary chloroplast (making a secondary chloroplast).[32]
Red algal derived chloroplasts
Cryptophytes
Cryptophytes, or cryptomonads are a group of algae that contain a red-algal derived chloroplast. Cryptophyte chloroplasts contain a nucleomorph that superficially resembles that of the chlorarachniophytes.[16] Cryptophyte chloroplasts have four membranes, the outermost of which is continuous with the rough endoplasmic reticulum. They synthesize ordinary starch, which is stored in granules found in the periplastid space—outside the original double membrane, in the place that corresponds to the red alga's cytoplasm. Inside cryptophyte chloroplasts is a pyrenoid and thylakoids in stacks of two.[14]
Their chloroplasts do not have phycobilisomes,[14] but they do have phycobilin pigments which they keep in their thylakoid space, rather than anchored on the outside of their thylakoid membranes.[14][16]
Cryptophytes may have played a key role in the spreading of red algal based chloroplasts.[47][48]
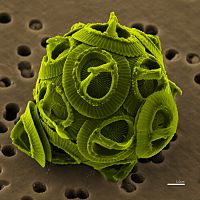
Scanning electron micrograph of Gephyrocapsa oceanica, a haptophyte.
Haptophytes
Haptophytes are similar and closely related to cryptophytes or heterokontophytes.[32] Their chloroplasts lack a nucleomorph,[14][16] their thylakoids are in stacks of three, and they synthesize chrysolaminarin sugar, which they store completely outside of the chloroplast, in the cytoplasm of the haptophyte.[14]
Heterokontophytes (stramenopiles)
The photosynthetic pigments present in their chloroplasts give diatoms a greenish-brown color.
The heterokontophytes, also known as the stramenopiles, are a very large and diverse group of eukaryotes. The photoautotrophic lineage, Ochrophyta, including the diatoms and the brown algae, golden algae,[33] and yellow-green algae, also contains red algal derived chloroplasts.[32]
Heterokont chloroplasts are very similar to haptophyte chloroplasts, containing a pyrenoid, triplet thylakoids, and with some exceptions,[14] having four layer plastidic envelope, the outermost epiplastid membrane connected to the endoplasmic reticulum. Like haptophytes, heterokontophytes store sugar in chrysolaminarin granules in the cytoplasm.[14] Heterokontophyte chloroplasts contain chlorophyll a and with a few exceptions[14]chlorophyll c,[16] but also have carotenoids which give them their many colors.[33]
Cyanobacteria
Archæplastida
Land plants
Glaucophyta
Green algae
Excavata
Euglenophyta
Rhodophyta
Chromalveolata
Rhizaria a
Paulinella
Chlorarachniophyta
Haptophyta
Cryptophyta
Heterokontophyta
Dinoflagellata
Apicomplexa
Ciliatea
Possible cladogram of chloroplast evolution[16][17][32] Circles represent endosymbiotic events. For clarity, dinophyte tertiary endosymbioses and many nonphotosynthetic lineages have been omitted.
a It is now established that Chromalveolata is paraphyletic to Rhizaria.[32]
Edit
Apicomplexans, chromerids, and dinophytes
The alveolates are a major clade of unicellular eukaryotes of both autotrophic and heterotrophic members. The most notable shared characteristic is the presence of cortical (outer-region) alveoli (sacs). These are flattened vesicles (sacs) packed into a continuous layer just under the membrane and supporting it, typically forming a flexible pellicle (thin skin). In dinoflagellates they often form armor plates. Many members contain a red-algal derived plastid. One notable characteristic of this diverse group is the frequent loss of photosynthesis. However, a majority of these heterotrophs continue to process a non-photosynthetic plastid.[49]
Apicomplexans
Apicomplexans are a group of alveolates. Like the helicosproidia, they're parasitic, and have a nonphotosynthetic chloroplast.[32] They were once thought to be related to the helicosproidia, but it is now known that the helicosproida are green algae rather than part of the CASH lineage.[32] The apicomplexans include Plasmodium, the malaria parasite. Many apicomplexans keep a vestigial red algal derived chloroplast[50][32] called an apicoplast, which they inherited from their ancestors. Other apicomplexans like Cryptosporidium have lost the chloroplast completely.[50] Apicomplexans store their energy in amylopectin granules that are located in their cytoplasm, even though they are nonphotosynthetic.[14]
Apicoplasts have lost all photosynthetic function, and contain no photosynthetic pigments or true thylakoids. They are bounded by four membranes, but the membranes are not connected to the endoplasmic reticulum.[14] The fact that apicomplexans still keep their nonphotosynthetic chloroplast around demonstrates how the chloroplast carries out important functions other than photosynthesis. Plant chloroplasts provide plant cells with many important things besides sugar, and apicoplasts are no different—they synthesize fatty acids, isopentenyl pyrophosphate, iron-sulfur clusters, and carry out part of the heme pathway.[50] This makes the apicoplast an attractive target for drugs to cure apicomplexan-related diseases.[30] The most important apicoplast function is isopentenyl pyrophosphate synthesis—in fact, apicomplexans die when something interferes with this apicoplast function, and when apicomplexans are grown in an isopentenyl pyrophosphate-rich medium, they dump the organelle.[50]
Chromerids
The Chromerida is a newly discovered group of algae from Australian corals which comprises some close photosynthetic relatives of the apicomplexans. The first member, Chromera velia, was discovered and first isolated in 2001. The discovery of Chromera velia with similar structure to the apicomplexanss, provides an important link in the evolutionary history of the apicomplexans and dinophytes. Their plastids have four membranes, lack chlorophyll c and use the type II form of RuBisCO obtained from a horizontal transfer event.[51]
Dinophytes
The dinoflagellates are yet another very large and diverse group of protists, around half of which are (at least partially) photosynthetic.[33][46]
Most dinophyte chloroplasts are secondary red algal derived chloroplasts. Many other dinophytes have lost the chloroplast (becoming the nonphotosynthetic kind of dinoflagellate), or replaced it though tertiary endosymbiosis[52]—the engulfment of another eukaryotic algae containing a red algal derived chloroplast. Others replaced their original chloroplast with a green algal derived one.[16][32][46]
Most dinophyte chloroplasts contain form II RuBisCO, at least the photosynthetic pigments chlorophyll a, chlorophyll c2, beta-carotene, and at least one dinophyte-unique xanthophyll (peridinin, dinoxanthin, or diadinoxanthin), giving many a golden-brown color.[49][46] All dinophytes store starch in their cytoplasm, and most have chloroplasts with thylakoids arranged in stacks of three.[14]
Ceratium furca, a peridinin-containing dinophyte[53]
The most common dinophyte chloroplast is the peridinin-type chloroplast, characterized by the carotenoid pigment peridinin in their chloroplasts, along with chlorophyll a and chlorophyll c2.[16][46] Peridinin is not found in any other group of chloroplasts.[46] The peridinin chloroplast is bounded by three membranes (occasionally two),[14] having lost the red algal endosymbiont's original cell membrane.[16][32] The outermost membrane is not connected to the endoplasmic reticulum.[14][46] They contain a pyrenoid, and have triplet-stacked thylakoids. Starch is found outside the chloroplast.[14] An important feature of these chloroplasts is that their chloroplast DNA is highly reduced and fragmented into many small circles. Most of the genome has migrated to the nucleus, and only critical photosynthesis-related genes remain in the chloroplast.[46]
The peridinin chloroplast is thought to be the dinophytes' "original" chloroplast,[46] which has been lost, reduced, replaced, or has company in several other dinophyte lineages.[32]
Karenia brevis is a fucoxanthin-containing dynophyte responsible for algal blooms called "red tides".[46]
The fucoxanthin dinophyte lineages (including Karlodinium and Karenia)[32] lost their original red algal derived chloroplast, and replaced it with a new chloroplast derived from a haptophyte endosymbiont. Karlodinium and Karenia probably took up different heterokontophytes.[32] Because the haptophyte chloroplast has four membranes, tertiary endosymbiosis would be expected to create a six membraned chloroplast, adding the haptophyte's cell membrane and the dinophyte's phagosomal vacuole.[54] However, the haptophyte was heavily reduced, stripped of a few membranes and its nucleus, leaving only its chloroplast (with its original double membrane), and possibly one or two additional membranes around it.[32][54]
Fucoxanthin-containing chloroplasts are characterized by having the pigment fucoxanthin (actually 19′-hexanoyloxy-fucoxanthin and/or 19′-butanoyloxy-fucoxanthin) and no peridinin. Fucoxanthin is also found in haptophyte chloroplasts, providing evidence of ancestry.[46]
Dinophysis acuminata has chloroplasts taken from a cryptophyte.[16]
Diatom-derived dinophyte chloroplasts
Some dinophytes, like Kryptoperidinium and Durinskia[32] have a diatom (heterokontophyte) derived chloroplast.[16] These chloroplasts are bounded by up to five membranes,[16] (depending on whether you count the entire diatom endosymbiont as the chloroplast, or just the red algal derived chloroplast inside it). The diatom endosymbiont has been reduced relatively little—it still retains its original mitochondria,[32] and has endoplasmic reticulum, ribosomes, a nucleus, and of course, red algal derived chloroplasts—practically a complete cell,[55] all inside the host's endoplasmic reticulum lumen.[32] However the diatom endosymbiont can't store its own food—its storage polysaccharide is found in granules in the dinophyte host's cytoplasm instead.[14][55] The diatom endosymbiont's nucleus is present, but it probably can't be called a nucleomorph because it shows no sign of genome reduction, and might have even been expanded.[32] Diatoms have been engulfed by dinoflagellates at least three times.[32]
The diatom endosymbiont is bounded by a single membrane,[46] inside it are chloroplasts with four membranes. Like the diatom endosymbiont's diatom ancestor, the chloroplasts have triplet thylakoids and pyrenoids.[55]
In some of these genera, the diatom endosymbiont's chloroplasts aren't the only chloroplasts in the dinophyte. The original three-membraned peridinin chloroplast is still around, converted to an eyespot.[16][32]
Kleptoplastidy
Main article: Kleptoplastidy
In some groups of mixotrophic protists, like some dinoflagellates (e.g. Dinophysis), chloroplasts are separated from a captured alga and used temporarily. These klepto chloroplasts may only have a lifetime of a few days and are then replaced.[56][57]
Cryptophyte-derived dinophyte chloroplast
Members of the genus Dinophysis have a phycobilin-containing[54] chloroplast taken from a cryptophyte.[16] However, the cryptophyte is not an endosymbiont—only the chloroplast seems to have been taken, and the chloroplast has been stripped of its nucleomorph and outermost two membranes, leaving just a two-membraned chloroplast. Cryptophyte chloroplasts require their nucleomorph to maintain themselves, and Dinophysis species grown in cell culture alone cannot survive, so it is possible (but not confirmed) that the Dinophysis chloroplast is a kleptoplast—if so, Dinophysis chloroplasts wear out and Dinophysis species must continually engulf cryptophytes to obtain new chloroplasts to replace the old ones.[46]
Chloroplast DNA
Main article: Chloroplast DNA
See also: List of sequenced plastomes
Chloroplasts have their own DNA,[58] often abbreviated as ctDNA,[59] or cpDNA.[60] It is also known as the plastome. Its existence was first proved in 1962,[40] and first sequenced in 1986—when two Japanese research teams sequenced the chloroplast DNA of liverwort and tobacco.[61] Since then, hundreds of chloroplast DNAs from various species have been sequenced, but they are mostly those of land plants and green algae—glaucophytes, red algae, and other algal groups are extremely underrepresented, potentially introducing some bias in views of "typical" chloroplast DNA structure and content.[62]
Molecular structure
cytochrome
photosystem I
acetyl-CoA carboxylase
rubisco
tRNAs
tRNA
photosystem II
tRNAs
tRNAs
photosystem II
ribosomal proteins
tRNA
tRNA
nadh dehydrogenase
ribosomal proteins
tRNA
replication origin regions
tRNA
small RNA
ribosomal protein
replication origin regions
ribosomal RNA
tRNAs
ribosomal RNA
tRNA
cytochromes
photosystem II
ribosomal proteins
photosystem I
cytochromes
photosystem II
atp synthase
tRNAs
nadh dehydrogenase
tRNA
ribosomal proteins
photosystem I
tRNAs
photosystem II
RNA polymerase
ribosomal protein
atp synthase
tRNAs
ribosomal protein
tRNA
photosystem II
tRNA
tRNA
ribosomal RNA
tRNA
ribosomal RNA
tRNA
ribosomal protein
photosystem I
nadh dehydrogenase
tRNA
ribosomal protein
nadh dehydrogenase
tRNA
tRNA
ribosomal proteins
initiation factor 1
ribosomal proteins
RNA polymerase
atp-dependent protease
ribosomal proteins
tRNAs
nicotiana tabacum
edit · image
Chloroplast DNA Interactive gene map of chloroplast DNA from Nicotiana tabacum. Segments with labels on the inside reside on the B strand of DNA, segments with labels on the outside are on the A strand. Notches indicate introns.
With few exceptions, most chloroplasts have their entire chloroplast genome combined into a single large circular DNA molecule,[62] typically 120,000–170,000 base pairs long.[40][41][42][18] They can have a contour length of around 30–60 micrometers, and have a mass of about 80–130 million daltons.[63]
While usually thought of as a circular molecule, there is some evidence that chloroplast DNA molecules more often take on a linear shape.[62][64]
Inverted repeats
Many chloroplast DNAs contain two inverted repeats, which separate a long single copy section (LSC) from a short single copy section (SSC).[42] While a given pair of inverted repeats are rarely completely identical, they are always very similar to each other, apparently resulting from concerted evolution.[62]
The inverted repeats vary wildly in length, ranging from 4,000 to 25,000 base pairs long each and containing as few as four or as many as over 150 genes.[62] Inverted repeats in plants tend to be at the upper end of this range, each being 20,000–25,000 base pairs long.[42][65]
The inverted repeat regions are highly conserved among land plants, and accumulate few mutations.[42][65] Similar inverted repeats exist in the genomes of cyanobacteria and the other two chloroplast lineages (glaucophyta and rhodophyceae), suggesting that they predate the chloroplast,[62] though some chloroplast DNAs have since lost[65][66] or flipped the inverted repeats (making them direct repeats).[62] It is possible that the inverted repeats help stabilize the rest of the chloroplast genome, as chloroplast DNAs which have lost some of the inverted repeat segments tend to get rearranged more.[66]
Nucleoids
New chloroplasts may contain up to 100 copies of their DNA,[40] though the number of chloroplast DNA copies decreases to about 15–20 as the chloroplasts age.[67] They are usually packed into nucleoids, which can contain several identical chloroplast DNA rings. Many nucleoids can be found in each chloroplast.[63] In primitive red algae, the chloroplast DNA nucleoids are clustered in the center of the chloroplast, while in green plants and green algae, the nucleoids are dispersed throughout the stroma.[68]
Though chloroplast DNA is not associated with true histones,[9] in red algae, similar proteins that tightly pack each chloroplast DNA ring into a nucleoid have been found.[68]
DNA repair
In chloroplasts of the moss Physcomitrella patens, the DNA mismatch repair protein Msh1 interacts with the recombinational repair proteins RecA and RecG to maintain chloroplast genome stability.[69] In chloroplasts of the plant Arabidopsis thaliana the RecA protein maintains the integrity of the chloroplast’s DNA by a process that likely involves the recombinational repair of DNA damage.[70]
DNA replication
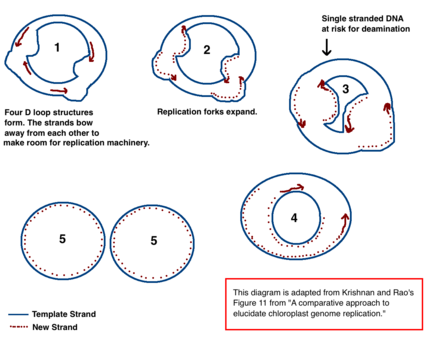
The leading model of cpDNA replication
Chloroplast DNA replication via multiple D loop mechanisms. Adapted from Krishnan NM, Rao BJ's paper "A comparative approach to elucidate chloroplast genome replication."
The mechanism for chloroplast DNA (cpDNA) replication has not been conclusively determined, but two main models have been proposed. Scientists have attempted to observe chloroplast replication via electron microscopy since the 1970s.[71][72] The results of the microscopy experiments led to the idea that chloroplast DNA replicates using a double displacement loop (D-loop). As the D-loop moves through the circular DNA, it adopts a theta intermediary form, also known as a Cairns replication intermediate, and completes replication with a rolling circle mechanism.[71][73] Transcription starts at specific points of origin. Multiple replication forks open up, allowing replication machinery to transcribe the DNA. As replication continues, the forks grow and eventually converge. The new cpDNA structures separate, creating daughter cpDNA chromosomes.
In addition to the early microscopy experiments, this model is also supported by the amounts of deamination seen in cpDNA.[71] Deamination occurs when an amino group is lost and is a mutation that often results in base changes. When adenine is deaminated, it becomes hypoxanthine. Hypoxanthine can bind to cytosine, and when the XC base pair is replicated, it becomes a GC (thus, an A → G base change).[74]
Over time, base changes in the DNA sequence can arise from deamination mutations. When adenine is deaminated, it becomes hypoxanthine, which can pair with cytosine. During replication, the cytosine will pair with guanine, causing an A --> G base change.
Deamination
In cpDNA, there are several A → G deamination gradients. DNA becomes susceptible to deamination events when it is single stranded. When replication forks form, the strand not being copied is single stranded, and thus at risk for A → G deamination. Therefore, gradients in deamination indicate that replication forks were most likely present and the direction that they initially opened (the highest gradient is most likely nearest the start site because it was single stranded for the longest amount of time).[71] This mechanism is still the leading theory today; however, a second theory suggests that most cpDNA is actually linear and replicates through homologous recombination. It further contends that only a minority of the genetic material is kept in circular chromosomes while the rest is in branched, linear, or other complex structures.[71][73]
Alternative model of replication
One of competing model for cpDNA replication asserts that most cpDNA is linear and participates in homologous recombination and replication structures similar to the linear and circular DNA structures of bacteriophage T4.[73][75] It has been established that some plants have linear cpDNA, such as maize, and that more species still contain complex structures that scientists do not yet understand.[73] When the original experiments on cpDNA were performed, scientists did notice linear structures; however, they attributed these linear forms to broken circles.[73] If the branched and complex structures seen in cpDNA experiments are real and not artifacts of concatenated circular DNA or broken circles, then a D-loop mechanism of replication is insufficient to explain how those structures would replicate.[73] At the same time, homologous recombination does not expand the multiple A --> G gradients seen in plastomes.[71] Because of the failure to explain the deamination gradient as well as the numerous plant species that have been shown to have circular cpDNA, the predominant theory continues to hold that most cpDNA is circular and most likely replicates via a D loop mechanism.
Gene content and protein synthesis
The chloroplast genome most commonly includes around 100 genes[24][41] that code for a variety of things, mostly to do with the protein pipeline and photosynthesis. As in prokaryotes, genes in chloroplast DNA are organized into operons.[24] Unlike prokaryotic DNA molecules, chloroplast DNA molecules contain introns (plant mitochondrial DNAs do too, but not human mtDNAs).[76]
Among land plants, the contents of the chloroplast genome are fairly similar.[42]
Chloroplast genome reduction and gene transfer
Over time, many parts of the chloroplast genome were transferred to the nuclear genome of the host,[40][41][77] a process called endosymbiotic gene transfer. As a result, the chloroplast genome is heavily reduced compared to that of free-living cyanobacteria. Chloroplasts may contain 60–100 genes whereas cyanobacteria often have more than 1500 genes in their genome.[78] Recently, a plastid without a genome was found, demonstrating chloroplasts can lose their genome during endosymbiotic the gene transfer process.[79]
Endosymbiotic gene transfer is how we know about the lost chloroplasts in many CASH lineages. Even if a chloroplast is eventually lost, the genes it donated to the former host's nucleus persist, providing evidence for the lost chloroplast's existence. For example, while diatoms (a heterokontophyte) now have a red algal derived chloroplast, the presence of many green algal genes in the diatom nucleus provide evidence that the diatom ancestor had a green algal derived chloroplast at some point, which was subsequently replaced by the red chloroplast.[44]
In land plants, some 11–14% of the DNA in their nuclei can be traced back to the chloroplast,[39] up to 18% in Arabidopsis, corresponding to about 4,500 protein-coding genes.[80] There have been a few recent transfers of genes from the chloroplast DNA to the nuclear genome in land plants.[41]
Of the approximately 3000 proteins found in chloroplasts, some 95% of them are encoded by nuclear genes. Many of the chloroplast's protein complexes consist of subunits from both the chloroplast genome and the host's nuclear genome. As a result, protein synthesis must be coordinated between the chloroplast and the nucleus. The chloroplast is mostly under nuclear control, though chloroplasts can also give out signals regulating gene expression in the nucleus, called retrograde signaling.[81]
Protein synthesis
See also: Transcription and translation
Protein synthesis within chloroplasts relies on two RNA polymerases. One is coded by the chloroplast DNA, the other is of nuclear origin. The two RNA polymerases may recognize and bind to different kinds of promoters within the chloroplast genome.[82] The ribosomes in chloroplasts are similar to bacterial ribosomes.[83]
This section needs expansion with: Genome size differences between algae and land plants, chloroplast stuff coded by the nucleus. You can help by adding to it. (January 2013)
Protein targeting and import
See also: Translation
Because so many chloroplast genes have been moved to the nucleus, many proteins that would originally have been translated in the chloroplast are now synthesized in the cytoplasm of the plant cell. These proteins must be directed back to the chloroplast, and imported through at least two chloroplast membranes.[84]
Curiously, around half of the protein products of transferred genes aren't even targeted back to the chloroplast. Many became exaptations, taking on new functions like participating in cell division, protein routing, and even disease resistance. A few chloroplast genes found new homes in the mitochondrial genome—most became nonfunctional pseudogenes, though a few tRNA genes still work in the mitochondrion.[78] Some transferred chloroplast DNA protein products get directed to the secretory pathway[78] though it should be noted that many secondary plastids are bounded by an outermost membrane derived from the host's cell membrane, and therefore topologically outside of the cell, because to reach the chloroplast from the cytosol, you have to cross the cell membrane, just like if you were headed for the extracellular space. In those cases, chloroplast-targeted proteins do initially travel along the secretory pathway.[32]
Because the cell acquiring a chloroplast already had mitochondria (and peroxisomes, and a cell membrane for secretion), the new chloroplast host had to develop a unique protein targeting system to avoid having chloroplast proteins being sent to the wrong organelle.[84]
The two ends of a polypeptide are called the N-terminus, or amino end, and the C-terminus, or carboxyl end.[85] This polypeptide has four amino acids linked together. At the left is the N-terminus, with its amino (H2N) group in green. The blue C-terminus, with its carboxyl group (CO2H) is at the right.
In most, but not all cases, nuclear-encoded chloroplast proteins are translated with a cleavable transit peptide that's added to the N-terminus of the protein precursor. Sometimes the transit sequence is found on the C-terminus of the protein,[86] or within the functional part of the protein.[84]
Transport proteins and membrane translocons
After a chloroplast polypeptide is synthesized on a ribosome in the cytosol, an enzyme specific to chloroplast proteins[87]phosphorylates, or adds a phosphate group to many (but not all) of them in their transit sequences.[84] Phosphorylation helps many proteins bind the polypeptide, keeping it from folding prematurely.[84] This is important because it prevents chloroplast proteins from assuming their active form and carrying out their chloroplast functions in the wrong place—the cytosol.[88][89] At the same time, they have to keep just enough shape so that they can be recognized by the chloroplast.[88] These proteins also help the polypeptide get imported into the chloroplast.[84]
From here, chloroplast proteins bound for the stroma must pass through two protein complexes—the TOC complex, or translocon on the outer chloroplast membrane, and the TIC translocon, or translocon on the inner chloroplast membrane translocon.[84] Chloroplast polypeptide chains probably often travel through the two complexes at the same time, but the TIC complex can also retrieve preproteins lost in the intermembrane space.[84]
Structure
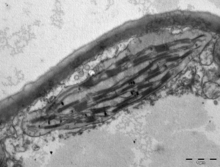
Transmission electron microscope image of a chloroplast. Grana of thylakoids and their connecting lamellae are clearly visible.
In land plants, chloroplasts are generally lens-shaped, 3–10 μm in diameter and 1–3 μm thick.[90][18] Corn seedling chloroplasts are ≈20 µm3 in volume.[18] Greater diversity in chloroplast shapes exists among the algae, which often contain a single chloroplast[14] that can be shaped like a net (e.g., Oedogonium),[91] a cup (e.g., Chlamydomonas),[92] a ribbon-like spiral around the edges of the cell (e.g., Spirogyra),[93] or slightly twisted bands at the cell edges (e.g., Sirogonium).[94] Some algae have two chloroplasts in each cell; they are star-shaped in Zygnema,[95] or may follow the shape of half the cell in order Desmidiales.[96] In some algae, the chloroplast takes up most of the cell, with pockets for the nucleus and other organelles,[14] for example, some species of Chlorella have a cup-shaped chloroplast that occupies much of the cell.[97]
All chloroplasts have at least three membrane systems—the outer chloroplast membrane, the inner chloroplast membrane, and the thylakoid system. Chloroplasts that are the product of secondary endosymbiosis may have additional membranes surrounding these three.[43] Inside the outer and inner chloroplast membranes is the chloroplast stroma, a semi-gel-like fluid[30] that makes up much of a chloroplast's volume, and in which the thylakoid system floats.
437 132 How do I fold or collapse sections of code in Visual Studio Code? Is this feature supported? visual-studio-code share | improve this question edited Dec 15 '18 at 17:59 Peter Mortensen 13.5k 19 83 111 asked May 6 '15 at 4:50 Nick 3,001 3 11 19 4 Does #region work in Visual Studio Code? The code folding feature isn't supported, but maybe that's an alternative. – Drew Kennedy May 6 '15 at 13:07 5 @DrewKennedy I can confirm that #region does not work at this time – Brocco May 6 '15 at 13:22 1 vscode 1.17 (September 2017) introduced region code.visualstudio.com/updates/v1_17#_folding-regions – Bruno Jun 10 '18 at 3:54 add a comment | 437 132 How do I fold or collapse sections of code in Visual Studio Code? Is this feature supported? visual-studio-code share | improve this question edited Dec 15 '18 at 17:59...


![Primary endosymbiosis A eukaryote with mitochondria engulfed a cyanobacterium in an event of serial primary endosymbiosis, creating a lineage of cells with both organelles.[9] It is important to note that the cyanobacterial endosymbiont already had a double membrane—the phagosomal vacuole-derived membrane was lost.[16]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Chloroplast_endosymbiosis_simple.svg/540px-Chloroplast_endosymbiosis_simple.svg.png)

















![The two ends of a polypeptide are called the N-terminus, or amino end, and the C-terminus, or carboxyl end.[85] This polypeptide has four amino acids linked together. At the left is the N-terminus, with its amino (H2N) group in green. The blue C-terminus, with its carboxyl group (CO2H) is at the right.](http://upload.wikimedia.org/wikipedia/commons/thumb/c/c0/Tetrapeptide_structural_formulae.svg/350px-Tetrapeptide_structural_formulae.svg.png)